To view an animation of the bacterial flagellum, click here.
The bacterial flagellum is an example of what Michael Behe describes as an irreducibly complex system. In his book, Darwin's Black Box, he explains that such irreducibly complex systems could not have arisen by a gradual step-by-step Darwinian process.
Because the bacterial flagellum is necessarily composed of at least three parts -- a paddle,a rotor, and a motor -- it is irreducibly complex. Gradual evolution of the flagellum, like the cilium, therefore faces mammoth hurdles. (p. 72)
Behe summarizes the structure of the bacterial flagellum in these terms:
Some bacteria boast a marvelous swimming device, the flagellum, which has no counterpart in more complex cells. In 1973 it was discovered that some bacteria swim by rotating their flagella. So the bacterial flagellum acts as a rotary propellor -- in contrast to the cilium, which acts more like an oar.
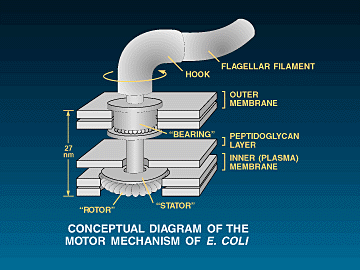
The structure of a flagellum is quite different from that of a cilium. The flagellum is a long, hairlike filament embedded in the cell membrane. The external filament consists of a single type of protein, called "flagellin." The flagellin filament is the paddle surface that contacts the the liquid during swimming. At the end of the flagellin filament near the surface of the cell, there is a bulge in the thickness of the flagellum. It is here that the filament attaches to the rotor drive. The attachment material is comprised of something called "hook protein." The filament of a bacterial flagellum, unlike a cilium, contains no motor protein; if it is broken off, the filament just floats stiffly in the water. Therefore the motor that rotates the filament-propellor must be located somewhere else. Experiments have demonstrated that it is located at the base of the flagellum, where electron microscopy shows several ring structures occur. The rotary nature of the flagellum has clear, unavoidable consequences ... (pp. 70-72)
The consequences Behe refers to are inferred by the nature of its irreducibly complex components, the discovery of which undermines a Darwinian explanation of origins. Behe concludes:
In summary, as biochemists have begun to examine apparently simple structures like cilia and flagella, they have discovered staggering complexity, with dozens or even hundreds of precisely tailored parts. It is very likely that many of the parts we have not considered here are required for any cilium to function in a cell. As the number of required parts increases, the difficulty of gradually putting the system together skyrockets, and the likelihood of indirect scenarios plummets. Darwin looks more and more forlorn. New research on the roles of the auxiliary proteins cannot simplify the irreducibly complex syetem The intransigence of the problem cannot be alleviated; it will only get worse. Darwinian theory has given no explanation for the cilium or flagellum. The overwhelming complexity of the swimming systems push us to think it may never give an explanation. (p. 73)
Behe concludes that such irreducibly complex systems were ultimately the result of intelligent design.
(It should be pointed out that Behe has no objections to the concept of universal common ancestry. His objections to evolution are limited to the rejection of the neo-Darwinian mechanism as a sufficient explanation for the origin of all biological systems.)
Most scientists will argue that "intelligent design" is not a purely naturalistic or materialistic explanation of the origin of biological information and hence is not a valid scientific explanation. Often hidden within this argument is the philosophical or theological assumption that God (or any other intelligent agent) did not play any active, direct, or discernable role in the creation of any biological system. This assumption requires that all genetic information must have arisen on a totally random and/or gradual step-by-step basis. It follows that nature must be continuous and natural history must be represented by a tree of life. Constrained by naturalism, the hypothesis of common ancestry must be accepted axiomatically and hence is not subject to scientific falsification.
Ironically, scientists are more constrained under a paradigm of naturalism than they are under a paradigm of intelligent design. Intelligent design of the universe may have included the creation of natural laws and initial conditions sufficient to allow the evolution of a single tree of life on earth. Intelligent design may also have included punctuations throughout natural history, direct acts of creation or the infusion of new genetic information, which resulted in natural discontinuities. Examples of such natural discontinuities might include the origin of life, the origin of the higher taxa (e.g. the origin of most of the major phyla in the Cambrian explosion), or the origin of irreducibly complex systems. If natural discontinuities exist, natural history would be more accurately modeled as a forest of life. The important point is that a paradigm of intelligent design can accommodate either a tree or forest of life as well as the existence of processes that either produce or prevent major evolutionary change. Unlike the paradigm of naturalism, however, intelligent design requires that scientists develop testable hypotheses to determine which model best fits the data.
The goal of science is to most accurately describe and model the Cosmos. While there may be circumstantial evidence to support either a continuous or discontinuous model of life, it is vitally important to recognize how one's starting assumptions affect one's conclusions. Beginning either with the assumption of intelligent design or philosophical agnosticism (both of which allow the Cosmos to be either continuous or discontinuous) clearly gives the scientist greater freedom of thought than exists within a purely naturalistic paradigm. Along with that freedom comes the responsibility to test what philosophical naturalists simply assume.
There are numerous examples of organisms with systems of highly specialized, interdependent components, all of which must be integrated before they are functional and offer any selective advantage. Nature abounds in such systems of "irreducible complexity." Although there may be some examples where certain components take on some preadaptive function, these cases may be considered as exceptions to a more general rule. As a rule, any subset of the components would prove to be a burden to an organism and thus be eliminated by natural selection. Take for example the sensory and motor mechanism of the common bacterium, Escherichia coli, a relatively simple unicellular prokaryotic organism.
The sensory and motor mechanism consists of a number of receptors which initially detect the concentrations of a variety of chemicals. Secondary components extract information from these sensors which in turn is used as input to a gradient sensing mechanism. The output of this mechanism is used to drive a set of constant torque proton-powered reversible rotary motors which transfer their energy through a microscopic drive train and propel helical flagella. This highly integrated system allows the bacterium to migrate at the rate of approximately ten body lengths per second. Dr. Robert Macnab of Yale University concluded a major 50 page review of this mechanism with these remarks:
As a final comment, one can only marvel at the intricacy in a simple bacterium, of the total motor and sensory system which has been the subject of this review and remark that our concept of evolution by selective advantage must surely be an oversimplification. What advantage could derive, for example, from a "preflagellum" (meaning a subset of its components), and yet what is the probability of "simultaneous" development of the organelle at a level where it becomes advantageous?
Macnab, R. (1978), "Bacterial Mobility and Chemotaxis: The Molecular Biology of a Behavioral System," CRC Critical Reviews in Biochemistry, vol. 5, issue 4, Dec., pp. 291-341
A good general introduction to flagella can be found in Voet, D. and Voet, J. G. (1995) Biochemistry, 2nd edition, John Wiley and Sons, New York, pp. 1259-1260.
Greater detail about the flagellar motor can be found in the following:
Schuster, S. C. and Khan, S. (1994) "The Bacterial Flagellar Motor," Annual Review of Biophysics and Biomolecular Structure, 23, 509-539; Caplan, S. R. and Kara-Ivanov, M. (1993) "The Bacterial Flagellar Motor," International Review of Cytology, 147, 97-164.