September 1, 2004
No matter how hard you try, it’s impossible to stop all of your muscles from working all at once.Ā Even if you have decided to do absolutely nothing, you still need your muscles to maintain your position, even if it’s just for lying in bed.Ā And within a few seconds, the respiratory center in your brain will stimulate your muscles of respiration to make you breathe.Ā You also have to remember that the heart is a muscle that contracts every second or so to maintain the flow of blood throughout the body.Ā And don’t forget the smooth muscle cells that surround the gastrointestinal tract and the blood vessels.Ā They’re always contracting and relaxing in order to allow these systems to function properly to maintain your existence here on earth.Ā For the purposes of this column, I will focus on skeletal muscle, which is under voluntary control of the nervous system, and the biomolecular basis for its ability to contract.Ā
Each skeletal muscle consists of many individual muscle fibers that are grouped together in bundles. The muscle fibers are held together by connective tissue.Ā This tissue also attaches the muscle to bones by way of tendons which then enables the contracting muscle to transmit a mechanical force that will allow for action.
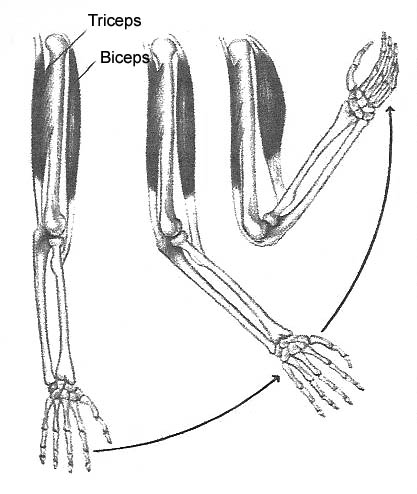
For example: when asked to show how strong they are, most people will raise their arms up so that they are about 90 degrees to their shoulders, and then forcefully bring their clenched fists up to their shoulders. In order to be able to accomplish this, their biceps muscles in each arm will have to maximally contract thereby forming a bulge, the size of which is used by many to judge someone’s overall strength.Ā The biceps muscle is able to do this because its origin tendon is located in the front part of the shoulder and its insertion tendon is located on the inner aspect of the forearm.Ā So when the biceps contracts, the elbow flexes.(see Figure 1)
Figure 1. Muscles function by contracting. The biceps muscle originates in the front part of the shoulder and its tendon attaches in the front part of the forearm. When the biceps contracts this lifts the forearm up towards the shoulder, i.e. the elbow flexes. The triceps muscle originates in the back part of the shoulder and its tendon attaches on the back part of the elbow. When it contracts this pulls the forearm down and away from the shoulder, i.e. the elbow extends.
In a similar manner, the triceps muscle is located on the back part of the upper arm, where it too has its origin in the shoulder, but not in front, but in the back, and its insertion is located on the back part of the elbow.Ā When the triceps contracts it will do the opposite of what the biceps does; namely, extend, or straighten out the elbow. (see Figure 1)Ā
One thing to keep in mind is that when you decide that you want to flex your biceps muscles to show how strong you are, the nervous system not only has to tell it to contract, but also the triceps muscles to relax, otherwise they would be working against each other.Ā How the nervous system accomplishes this will be answered in a future column, but I just wanted you to be aware of another level of complexity that is inherent in neuromuscular function.Ā It’s not as simple as it looks!Ā Before we look at how a muscle functions, let’s review how the nervous system is able to pass a message to the muscle to make it contract.ĀĀĀ
ReviewĀ
We are now at the point where the muscle cell’s end-plate has had its resting membrane potential depolarized due to the release of acetylcholine from the pre-synaptic motor neuron.Ā How do we get from this point to muscle cell contraction?Ā That’s what this column is all about.Ā I suspect that your wonder quotient may experience a significant
upturn when you’ve absorbed and seriously considered how this is all accomplished in the body.Ā Along the way, keep asking yourself how this could have all come into existence in a step by step fashion as macroevolution predicts.
Muscle Cell Structure
Each skeletal muscle fiber is a single cell that is long and cylindrical
in shape.Ā It is surrounded by a plasma membrane called the sarcolemma and it
contains numerous nuclei and two other structures calledĀ T-tubules (transverse
tubules) and the sarco-plasmic reticulum, both of which play an important role
in muscle contraction.Ā Within each of these muscle fibers are bundles of myofibrils
each of which is made up of repeating units called sarcomeres.Ā The sarcomere
consists of the contractile proteins and is the basic unit of the muscle cell.
Ā
These contractile proteins are called actin and myosin.Ā The actin consists of very thin filaments and the myosin is composed of thick filaments.Ā Under polarized light and electron microscopy the region containing actin is lighter and is bisected by a dark line known as the Z line.Ā The region between two Z lines is designated as a sarcomere.Ā The myosin filaments are in the center of the sarcomere within what is called the A- band (see Figure 2).Ā As the figure clearly demonstrates, at rest there is an overlapping of the actin and myosin filaments.Ā

Figure 2. Each sarcomere is bounded by two Z lines. Within the sarcomere are the thick myosin filaments which make up the A Band. In the resting state, the actin filaments overlap with the myosin filaments. Notice the molecular configuration of actin with myosin in the bottom part of the figure. This will help you to visualize what happens when contraction takes place.
The current understanding of muscle contraction is known as the sliding filament theory.
In essence, under the right set of circumstances, and in the presence of molecularly derived energy, actin and myosin, by repeated chemical interactions and molecular positioning, are able to literally slide by each other, thereby causing contraction of the muscle cell.Ā This can occur because the actin molecule has myosin-binding sites which allow for a piece of the myosin filament to repetitively interact with the actin filament.Ā This contraction results in the Z lines coming closer together as actin and myosin slide past each other. (see Figure 3)

Figure 3. The Myosin head acts as a cross-bridge to the actin molecule and in the presence of molecules that release energy in the cell, it is able to move along the actin molecule from binding site to binding site as actin moves in the opposite direction..
Standing in the way of this interaction, and therefore also muscle contraction, are two proteins called tropomyosin and troponin.Ā Tropomyosin is a long protein that wraps itself around the actin filament and literally lies over the myosin-binding sites and therefore is physically capable of blocking myosin from binding to actin. ĀMeanwhile, the troponin molecule sits on top of tropomyosin fixing it in place, thereby helping the process along.Ā In order for actin and myosin to interact it looks like tropomyosin will have to be moved off of the actin molecule. (see Figure 4)ĀĀĀ

Figure 4. The upper panel shows the binding sites for myocin on the actin molecule by way of small black dots within the small white circles. Tropomyosin can clearly be seen obstructing these mysoin-binding sites, thereby preventing myosin from interacting with actin. The lower panel shows what happens in the presence of Ca++ ions flooding the sarcoplasm. The Ca++ ion binds to the CA++ binding receptor on the troponin molecule which causes it to literally lift tropomyosin off of the myosin binding sites of actin, thereby allowing myosin to interact with actin and muscle contraction to take place.
Remember I said “under the right set of circumstances” actin and myosin, in the presence of energy, are able to slide by each other in order to effect muscle cell contraction.Ā
Just imagine if this actin-myosin interaction were taking place all of the time without direct control from the nervous system.Ā Remember how you were able to flex your biceps to show how strong you were?Ā How would you be able to do it if at the same time the triceps was also contracting?Ā And that goes for every other muscle system in the body.Ā Think about it!Ā Every joint is able to move in more than one direction, and for each directional movement, there are at least two complementary muscles that cause flexion-extension, abduction-adduction, and inner and outer rotation.Ā Clearly, in order for our survival on earth we require that our body be able to have some measure of muscular control. ĀĀ
Tropomyosin and troponin essentially have a lock on actin-myosin interaction and therefore it would appear that muscle contraction is impossible.Ā Let’s see how the nervous system is able to unlock this molecular puzzle and allow for controlled muscle contraction and survival.
Muscle Cell Contraction
Don’t forget that we diverted from this explanation of muscle contraction
to describe the structure of the muscle cell after having brought a motor neuron
impulse to the myo-neural junctionĀ This resulted in the release of acetylcholine
and the depolarization of the muscle membrane end plate.Ā From here on the depolarization
wave is passed on by way ofĀ the T-tubules through the plasma membrane to the
sarcoplasmic reticulum.Ā This structure contains Ca++ ions and in response to
depolarization, Ca++ ion channels open within it allowing for the release of
Ca++ ions into the cytoplasm of the muscle cell.
Interestingly enough, the increase of Ca++ ions in the sarcoplasm allows Ca++ ions to bind to the troponin molecule. ĀIn doing so this forms what is called a troponin-Ca++ complex, which undergoes a conformational change resulting in it changing shape.Ā When this happens, it literally pulls tropomyosin off of the myosin-binding sites of the actin molecule.Ā This release allows actin and myosin to interact once more and muscle contraction takes place. (see Figure 5) ĀĀ

Figure 5. Top panel shows muscle cell at rest with actin and myosin overlapping and sarcoplasmic reticulum full of Ca++ ions (black dots). Depolarization of the muscle end plate travels along the transverse tubules (T-tubules) to the sarcoplasmic reticulum. In the second panel, voltage-gated Ca++ ion channels in the sarcoplasmic reticulum open, allowing Ca++ ions to flood into the cytoplasm of the muscle cell (sarcoplasm). This allows actin and nmyosin to slide by each other causing contraction and the bringing together of the Z lines. The third panel shows the actin of the Ca++ ion pump within the sarcoplasmic reticulum pumping Ca++ ions back in. This results in relaxation of the muscle as shown by the return of actin, myosin, and the Z lines to their resting state, as seen in the first panel..
However, this actin-myosin interaction is limited by the fact that the sarcoplasmic reticulum contains a Ca++ ion pump which soon begins to pump Ca++ ions back into itself.Ā This causes the Ca++ ion level to drop within the cytoplasm and the release of Ca++ ions from the troponin molecule.Ā When this happens, the troponin-Ca++ complex reverts to troponin and its previous shape thereby allowing tropomyosin to once again block the myosin-binding sites on the Āactin molecule.Ā The subsequent uncoupling of actin and myosin then results in muscle relaxation and the muscle cell returns to its resting position.(see Figure 5)Ā All is now ready for another nerve impulse to cause stimulation of the muscle cell and controlled contraction.
Pathophysiology of Muscle Disease and Dysfunction
Last month’s column discussed conditions that affect the neuromuscular
junction, thereby rendering muscle tissue as being incapable of proper function.Ā
Conditions such as; botulism, myasthenia gravis, toxic exposure to insecticides,
and poison-tipped arrows.
Reviewing the information provided above and in last month’s column should lead one to recognize that unless the motor neuron is capable of releasing adequate amounts of acetylcholine (the neurotransmitter for muscle tissue), the muscle membrane contains specific acetylcholine receptors that directly affect the right ion channels to allow for muscle membrane depolarization, and the acetylcholine can be broken down in a timely manner to allow for proper repetitive muscle activity, adequate neuromuscular function within the body would be impossible.
Now having seen what is further required for muscle contraction once the motor nerve has caused the depolarization of the muscle end plate, let’s look at just a few of the ways that this system can breakdown and malfunction.Ā Without proper muscular function, one can readily see how a multi-system organism with a complex body plan would have difficulty in the survival of the fittest.
Muscular dystrophy is an inherited disorder of muscle tissue resulting in progressive muscle weakness and degeneration.Ā Many of these disorders are due to a genetic defect involving the production of one of the proteins necessary for supporting the muscle membrane during muscle contraction.Ā Absence, or defective function, of any of these supportive proteins results in the muscle membrane allowing too many Ca++ ions into the muscle cell which triggers certain enzymes within the cell to cause damage to the muscle fibers.Ā Eventually this results in defective muscle function manifesting as weakness.Ā
Channelopathies are defects that occur within different ion channels of the muscle membrane or the sarcoplasmic reticulum resulting in muscle dysfunction manifesting as problems with muscle tone and periodic paralysis.Ā There are also congenital muscle diseases that are related to abnormal structure within the muscle cell which result in dysfunction and weakness.
Questions for Macroevolution
In reviewing the mechanics behind muscle contraction, one is automatically
struck by the interdependence of the various components that are necessary for
this to occur.Ā After depolarization of the muscle membrane end plate, the muscle
requires the combined effects of:
Each of these components must exist and be in proper working order for controlled muscle contraction to take place.Ā In addition, as is evidenced by the various muscle disorders, any defect that results in muscle dysfunction is likely to result in the organism not being able to survive. Ā
Macroevolution’s claim for a step by step development of such a complicated system means that at some point in the past all of these components did not exist together as a working unit.Ā One must ask oneself what each component would have been doing without any number of the others and how this might have been useful to an organism with what would appear to us to be an incomplete neuromuscular system.Ā ĀĀ
If a muscle cell contained the proper contractile proteins but not the T-tubule system and the sarcoplasmic reticulum with its Ca++ ion pump, then how did controlled muscular activity allowing for survival take place?ĀĀ And what about the contractile proteins? Which came first; actin or myosin?Ā What purpose did each serve without the other?Ā What about tropomyosin and troponin?Ā Without both of them, controlled muscular activity is impossible.Ā Which came into existence first and what was its biological function without the other?Ā ĀĀĀĀ
These are the sorts of questions that in my mind need to be put forward and answered for students who are currently studying biology and the theories of its origin.Ā Now that one can understand the complex nature of neuromuscular function, one is capable of asking appropriate questions of science.Ā Of course, what I have provided here is only the tip of the iceberg in the complexity of neuromuscular function.Ā A fuller understanding of how all of this works should give one pause when considering the validity of macroevolution.
Of course, underlying all of this is the realization that the information necessary to create this incredibly complicated network of functioning neuromuscular tissue is thought to be wrapped up in the nucleus of the zygote.Ā As embryonic growth continues, its primordial tissue will eventually differentiate into over 200 different cell types, that are together, capable of all of this and much more.Ā
Wonder of wonders; miracle of miracles; one has to seriously question whether life’s origin has come about in such a simple manner as macroevolution would have us believe.
Our knowledge of the irreducible complexity ofĀ biomolecular systems combined with the medical knowledge of how easy it is for them to breakdown and die would seem to scream in our ears; “No way!!”
The Future Awaits
Starting next month we’ll be looking at how the body is capable of
converting light waves into what we perceive as vision.Ā I’m sure that
you’ll find the study intriguing especially as it may apply to the theory
of macroevolution.Ā I hope that after reading these three upcoming columns you’ll
come to the realization that; not only is it possible to be blind when the eyes
are capable of seeing, but also; when it comes right down to it, we really don’t
know how we are capable of experiencing the sensation of vision.Ā Does that
tweak your interest?Ā I hope so!!Ā Join me then in Part IV of Wired for Much
More Than Sound—Vision Part 1—The Parts of the Eye
Dr. G.
Howard Glicksman M. D. graduated from the University of Toronto in 1978. He practiced primary care medicine for almost 25 yrs in Oakville, Ontario and Spring Hill, Florida. He recently left his private practice and has started to practice palliative medicine for a Hospice organization in his community. He has a special interest in how the ethos of our culture has been influenced by modern science’s understanding and promotion of what it means to be a human being.
Copyright 2004 Dr. Howard Glicksman. All rights reserved. International
copyright secured.
File Date: 9.01.04